|
|||||||||||||||||||||||||||||||||||||||||||




 |
-
Primärproduzenten : Algen
-
Primärkonsumenten : Zooplankton, andere Herbivore
-
Sekundärkonsumenten: Carnivore
-
Endkonsumenten: Raubfische
-
Destruenten: u.a. Mückenlarven, Ringelwürmer, Pilze und Bakterien (Makroinvertebraten)
-
autolytische Abbauprozesse ( durch Enzyme Freisetzung zellularer Stoffe, die dann ins Wasser diffundieren)
Wie bereits erwähnt wird nur ein Teil der organischen Substanz bereits im Epilimninon zersetzt, während der weitaus größere Teil ins Hypolimnion gelangt, wo die Sauerstoffverhältnisse bestimmen, ob Oxidation der Reduktion als Abbauprozess in Frage kommen.
Sauerstoff und Kohlenstoffdioxid - die Schlüsselfaktoren im See
beeinflussende Faktoren:
-
Verbrauch und Produktion der Gase durch Organismen
-
physikalische Lösungs- und chemische Bindungsvermögen des Wassers unter verschiedenen Umweltbedingungen
O2 und CO2 sind gut wasserlöslich, die Löslichkeit ist jedoch abhängig von Temperatur und Druck. Mit steigender Temp. sinkt die Löslichkeit, wie auch bei abnehmenden Partialdrücken die Löslichkeit der Gase abnimmt.
Henry-Dalton'sches Gesetz: Die im Wasser gelösten Mengen der Gase sind ~ ihren Partialdrücken in der Atmosphäre.
Zirkulationsbewegungen: siehe Herbstzirkulation, Sommerstagnation
Stickstoffumsatz
Unter sauerstoffreichen Bedingungen werden Exkremente und Eiweißstoffe der abgestorbenen Lebewesen durch eiweißspaltende Pilze und Bakterien über eine Reihe von Zwischenstufen bis zu NH+4, CO2 und H2O abgebaut (Ammonifikation). Durch Bakterien der Gattung Nitrosomonas werden Ammonium-Ionen zu Nitrit- Ionen und durch die Gattung Nitrobacter Nitrit- zu Nitrat-Ionen oxidiert (Nitrifikation). Dabei wird die freigesetzte Energie von ihnen chemosynthetisch zur eigenen Versorgung mit organischen Substanzen genutzt. In nährstoffreichen Seen können die O2- verbrauchenden Abbauvorgänge bis zum völligen Aufzehren des O2 reichen, dann spricht man davon, dass der See umkippt. Dann laufen aerobe Reduktionsprozesse ab, die Nitrat-Ionen werden durch andere Bakteriengruppen wieder NH+4 (Nitratammonifikation) oder zu N2 (Denitrifikation).
Schwefelumsatz
-
hauptsächlich Sulfate (SO2- 4), organisch gebundener Schwefel (in Eiweißen) und H2S
-
neben biogenen Umsätzen, rein chem. Reaktionen, Oxidation von H2S durch O2 oder anaerobe Eisensulfidbildung im Sediment
Phosphorumsatz
- freigesetzte Phosphat-Ione aus organ. Stoffen mit O2 zu Fe (III)- Ionen gebunden
- Adsorption des entstehenden unlösliches Eisen(III)-phosphat (Fe(III)PO4 ) im Sediment
- bei anaeroben Bedingungen: Reduktion von Fe(III)PO4 zu Eisen (II)-phosphat
Mineralisierung
Umsetzung organischer Substanzen ( abgestorbene Organismen und Ausscheidungen) in mineralische Materie.
Endprodukte: CO2, NO3, SO2-4, Phosphat –Ione
Bei anaeroben Bedingungen entstehen: Schwefelwasserstoff (H2S), Ammoniak ( ) und Methan (
Bei fehlendem O2 nur teilweise Zersetzung, Bildung von Faulschlamm
|
|
oligotroph |
mesotroph |
eurotroph |
polytroph |
|
Pflanzennährstoffe |
+ |
++ |
+++ |
++++ |
|
Primärproduktion |
+ |
++ |
+++ |
++++ |
|
Sichttiefe |
++++ |
+++ |
++ |
+ |
|
Abbau organ. Stoffe |
++++ |
+++ |
++ |
+ |
|
Verlandungsprozess |
+ |
++ |
+++ |
++++ |
|
Artenreichtum |
++++ |
+++ |
++ |
+ |
|
Individuenanzahl |
+ |
++ |
+++ |
++++ |
Weiterhin spielt die Litoralausbildung und Sedimentation eine Rolle, je tiefer das Litoral reicht, desto das Produktionsprofil im Sommer und die Gasverteilung im Sommer spielen ebenfalls eine Rolle zur Klassifikation:
Initialstadium (vegetationsfreier Wasserkörper)
- Ansiedlung von planktischen Algen und Aufwuchsorganismen, Beginn einer geringen Primärproduktion
- Ansiedlung planktischer Konsumenten in einfachen Nahrungsketten
- Zuwachs an Arten und zunehmende Vernetzung von Trophieebenen, Stabilisierung eines Gleichgewichtes zwischen Auf- und Abbau
- langsame Nährstoffanreicherung
- allmähliches Überwiegen von Aufbauvorgängen gegenüber den Abbauprozessen unter dem Einfluss wachsender Nährstoff Angebote
- verstärkte Sedimentation durch unvollständigen Abbau organ. Substanz
- Verflachung und Verlandung
Klimaxstadium (Moor, Wald)
Nicht immer lassen sich alle einzelnen Vorgänge exakt beobachten, da die Sukzessionsverläufe von speziellen Bedingungen abhängig sind
Auch die Moorbildung richtet sich nach den Verhältnissen vor Ort. Nach dem Verlandungsprozess ist bald die gesamte Wasseroberfläche mit litoralen Pflanzen bedeckt. Nun kann sich ein Erlenbruch bilden. Der Abbau der großen Pflanzenmassen am Ende des Verlandungsstadiums kann zur Freisetzung großer Mengen CO2 und dem Sinken des pH- Wertes führen. Dann siedeln sich Azidophyten an. Unter solchen Bedingungen entstehen aus Flachmooren Graslandbiotope, die zunehmend trockener werden. dem Grasland folgen schließlich Gehölze.
In niederschlagsreichen Gebieten können sich in den versäuerten Flachmooren Torfmoose ansiedeln, die den pH- Wert weiter absenken. Die Torfmoose bilden dichte Polster die das Wasser halten. Wegen des niedrigen pH-Wertes können sich nur wenige abbauende Mikroorganismen hier etablieren. Das entstehende Hochmoor ist daher arm an wasserlöslichen Nährstoffen. Während die Torfmoose am oberen Teil ständig nachwachsen, zersetzen sich die abgestorbenen untere Teile nicht und in Folge dessen wächst der Körper weiter in die Höhe. Die abgestorbenen
Etwa 71% der Erdoberfläche werden vom Wasser bedeckt, somit stellen die Meere das größte zusammenhängende Ökosystem dar.
Dieser Lebensraum wird neben der Grobgliederung, Benthal und Pelagial, weiter horizontal und vertikal strukturiert. Nach der horizontalen Gliederung gibt es die küstennahen bis zu 200km breiten und 200m tiefen Schelfgebiete, an die sich der Kontinentalabhang anschließt, dem der Tiefseeboden folgt, der etwa fünf siebtel der Erdoberfläche ausmacht. Zum Litoral zählen der Küstenbereich und zur sublitoralen Zone gehören die tiefer gelegenen Bodenzonen und die Schelfgebiete.
Vertikal wird auch hier zwischen nach Lichteinfall unterschieden in eine euphotische und eine aphotische Zone.
- Salzgehalt (durchschnittl. 35 Promille)
- Nährstoffgehalt
- Temperatur
- Lichtverhältnisse
- Wasserbewegung
Im Sommer dagegen können die warmen Wassermassen über den kalten liegen und so eine Austauschbewegung verhindern (Sommerstagnation) In den Tropen herrschen also ganzjährig solche Verhältnisse oder anders die Wasseraustauschprozesse können nur in höheren Breiten stattfinden..

- marines Phytoplankton ( größtenteils aus Kieselalgen und Dinoflagelaten)
- Zooplankton (Krebstiere, Pfeilwürmer, Flügelschnecken und Larven von Krebsen, Ringelwürmern und Seeigeln)
- Nekton (sämtliche Wirbeltiere und Tintenfische)
Übrigens:
Süßwasserfische unterscheiden sich u.a. in ihren Trinkgewohnheiten von Meeresfischen. Während Meeresfische regelmäßig trinken, nehmen Süßwasserfiche niemals aktiv Wasser auf.
Der Grund dafür liegt in dem unterschiedlichem Salzgehalt des Wasser. Die schwächere Konzentration des Bluts gegenüber dem hohen Salzgehalt des Meerwassers führt zu einem ständigen Wasserverlust, wegen des osmotischen Gefälles. Als Ausgleich dafür müssen Meeresfische ca. 4-8% ihres Körpergewichtes an Flüssigkeit zu sich nehmen. Dem Trinkwasser wird zusätzlich über Kiemen und Darm Salz entzogen.
Süßwasserfische dagegen haben eine höhere Konzentration ihres Blutes gegenüber dem Wasser, was eine Einschwemmung von Wasser zur Folge hätte. Zum Ausgleich dafür nehmen sie kaum aktiv Wasser auf und scheiden über Haut und Urin bis zu 1/3 ihres Körpergewichtes aus. Salz-Ionen gewinnen sie über ihre Kiemen.
Konzentrationsschwankungen des Co2 und O2 - Gehaltes in Abhängigkeit zueinander
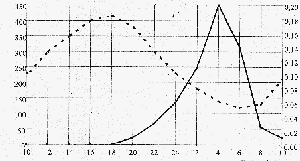
Gemessen wurde an einem sonnigen Julitag in einem Altlauf-Stillwasser bei einem pH-Wert von 7,5 -8 und einer Temperatur bei 20 -22°C. Die gestrichelte Linie stellt die O2 -Konzentration dar, die durchgehende Linie gibt den Gehalt an CO2 wieder.
Erkennbar werden ein Maximum des O2- Gehaltes bei 18.00Uhr und ein Maximum des Co2 Gehaltes bei 4.00Uhr, die Minima liegen für O2 bei 6.00Uhr und für CO2 bei 18.00 Uhr. Beide Stoffe unterliegen also einem tageszeitlich bedingten Zyklus.
Dabei fällt der Zusammenhang zwischen Maximal-Konzentration von Sauerstoff und gleichzeitiger Minimal-Konzentration an Kohlendioxid auf. Dies lässt sich dadurch erklären, dass Kohlenstoffdioxid durch fotoautotrophe Produzenten in Form von organischen Kohlenstoffverbindungen gebunden wird. Zur Vorbereitung der Kohlenstoffassimilation wird Wasser gespalten (Fotolyse), dadurch wird Sauerstoff frei. Da die erste Phase der Fotosynthese lichtabhängig ist, erreicht sie am Tage ihre Höchstleistung, während sie nachts rückgängig ist, sie setzt erst wieder ein nachdem der Lichtkompensationspunkt erreicht wurde.
Durch die Atmung wird andererseits Sauerstoff verbraucht und Kohlendioxid freigesetzt. Da nachts die lichtabhängige Phase der Fotosynthese nicht stattfinden kann, sinkt der Gehalt an O2 bei gleichzeitiger Zunahme von CO2.
Zwischen 4.00 und 6.00Uhr können wir ein gleichzeitiges Absinken von CO2 und O2 feststellen, was sich darauf zurückführen lässt, dass die Fotosynthese bereits wiedereingesetzt hat (Rückgang von CO2) aber das Licht noch nicht den erforderlichen Kompensationspunkt erreicht hat. Somit werden durch Dissimilationsvorgänge noch mehr O2 veratmet, als fotosynthetisch produziert wird. Erst wenn der Lichtkompensationspunkt überschritten wird, (an dem sich Dissimilation und Assimilation die Waage halten), nimmt die O2-Konzentration wieder zu.
Veränderungen durch sauren Regen
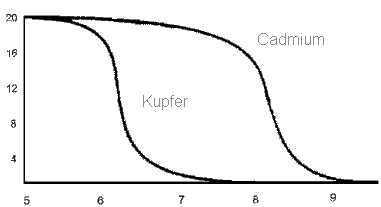
Metallgehalt eines Fließgewässers in Abhängigkeit vom pH-Wert (untere Leiste)
(nach: Kronberg, I., Natura -Ökologie. - Stuttgart, Klett, 1996, S.83)
Deutlich zu erkennen ist die Abhängigkeit der Schwermetallkonzentration vom pH-Werte, je niedriger der pH- Wert desto höher ist der Gehalt an Schwermetallen.
Saurer Regen enthält schweflige Säure und Schwefelsäure, bei einem solchen Eintrag in das Gewässer werden Schwermetall-Ionen aus dem Sediment gewaschen. Die Zunahme von Hydroxid -Ionen bewirkt eine Versauerung des Gewässers und führt zu dessen erhöhtem Schwermetallgehalt.
Die Giftwirkung der Schwermetalle auf den Organismus ist vor allem in ihrem Einfluss auf die Enzyme zu sehen. Eine mögliche Wirkung der Schwermetalle, könnte eine irreversible Schädigung der DNA-Polymerase sein. Dies hätte zur Folge, dass die DNA nicht mehr identisch repliziert werden kann und damit keine Mitose, also kein Wachstum mehr möglich ist.
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Seit einigen Jahren weiß man, dass auch bei Wasserpflanzen elektrophysiologische Prozesse zur Informationsübertragung vorkommen.
Die Wasserfalle, Aldrovande vesiculosa, eine nur bis 15 cm groß werdende fleischfressende Wasserpflanzen, kann wie die Venusfalle, durch Zusammenklappen ihrer Blättchen kleine Wassertiere fangen. Bei Berührung, z.B. durch ein Ruderfußkrebschen, werden die Aktionspotentiale ausgelöst.
Dabei ist es interessant diese pflanzlichen Aktionspotentiale mit denen menschlicher Neuriten zu vergleichen. Es fällt auf, dass die Dauer des Aktionspotentials bei der Pflanze sehr viel größer ist, die Nervenzelle eines Menschen bleibt nur 1-2 ms erregt bis sie in die Refraktärphase übergeht. Gleich ist jedoch bei beiden, dass vom Ruhepotential durch Depolarisation und nachfolgender Repolarisation der Aufbau eines Aktionspotentials erfolgt
Auch pflanzlichen Aktionspotentialen liegen Prozesse von Ionenbewegungen und unterschiedliche Ionenverteilungen an der Zellmembran zugrunde.
|
Ionensorte |
Membranaußenseite mmol/l |
Membraninnenseite mmol/l |
|
Cl- |
0,9 |
90,7 |
|
Mg2+ |
1,7 |
17,7 |
|
Ca2+ |
0,8 |
10,2 |
|
Na+ |
0,2 |
10,0 |
|
K+ |
0,04 |
54,3 |
|
Organische Anionen |
- |
reichlich vorhanden |
Ionenverteilung im Zellinneren und an der Membranaußenseite einer Wasserpflanze an einer nichterregten Membran, (us: Biologie-Unterrichtsmaterialien, Stark-Verlag, o. J.)
Die Ladungsverteilung gleicht grundsätzlich der am Neuriten. In der Membraninnenseite überwiegen negative Ladungsträger, vor allem organische Anionen Chlorid-Ionen sind etwa in hundertfacher, Kalium-Ionen in etwa tausendfacher Form vorhanden. Für das Aktionspotential sorgen die Ionen die in größter Ungleichverteilung vorliegen. Ein Aktionspotential kann sich also nur durch den Ausstrom von Chlorid-Ionen aufbauen, weil nur dadurch die Innenseite positiver werden kann. Die Redpolarisierung kann nur durch einen Ausstrom von Kalium-Ionen erfolgen, da die Innenseite nur dadurch wieder negativ werden kann. Da die anderen Kationen im Zellinnenraum in höherer Konzentration vorliegen, kann sich durch deren Einströmen kein Aktionspotential aufbauen. da der Einstrom nicht gegen ein Konzentrationsgefälle aufbauen kann.
Die Lebensgemeinschaft der Savanne
Zwischen dem Gürtel der subtropischen Wüsten und dem äquatorialen Regenwald liegen die Savannen.
Savannen sind Grasfluren, die von einzelnen Bäumen, Sträuchern oder Baumgruppen bestanden sind. Sie gehen ohne scharfe Grenzen in die Tropenwälder einerseits und in Steppen und Halbwüsten andererseits über.
Die entscheidenden abiotischen Faktoren sind
-
ganzjährlich hohe Temperaturen (20°C)
-
relativ hohe jährliche Niederschlagsmengen, mit einem deutlich ausgeprägtem Wechsel zwischen Trocken- und Regenzeit
-
episodische Feuer, die meist durch Gewitter, oft auch durch Menschen ausgelöst werden
wesentliche biotische Faktoren:
-
Gräser, sind die vorherrschenden Pflanzengruppe
-
Weidetiere, die in Herden das Gras abweiden und jahreszeitlich wandern
-
Raubtiere, die den Herden folgen und sich von ihnen ernähren
-
Dungkäfer, die die Exkremente der Weidetiere beseitigen
Gräser und Gehölze stehen miteinander im Wettbewerb, das Gleichgewicht wird durch Beweidung, episodische Feuer und periodische Dürren aufrechtgehalten.
Gräser und Gehölze sind Konkurrenten, wobei die Gehölze den Gräsern überlegen sind, da sie durch ihr kontinuierliches Höhenwachstum den Gräsern das Licht nehmen. Grasland kann nur dort überleben, wo die Gehölze keine Chance haben. In der Savanne gibt es drei Faktoren, die die Ausbreitung der Gehölze verhindern : Trockenheit, Feuer und Weidetiere. Gräser haben sich an diese Faktoren angepasst.
Gräser zeichnen sich durch Trockenresistenz aus. Die Blätter schützen sich dadurch, dass sie sich der Länge nach einrollen. Bei Dürre sterben die Halme oberflächlich ab und schützen als trockenen Hülle die dicht an der Erdoberfläche liegenden Sprossanlagen. Zur Regenzeit entfalten sie sich sehr schnell, blühen und fruchten nach kurzer Zeit. Durch ihr dichtes und flaches Wurzelwerk können sie Niederschläge rasch und effektiv aufnehmen.
Auch wenn das trockene Gras leicht entflammt sind Gräser resistent gegen Feuer. Seine Wachstumszonen liegen, vor der Hitze sicher, unterhalb der Erdoberfläche.
Dadurch dass sie schnell nachwachsen und Seitentriebe bilden, überleben Gräser den Abtritt durch die Herden ebenso gut wie das regelmäßige Abweiden.
Die Huftierherden nutzen den Nährwert des Grases.
Die Evolution der Weidetiere wird vom Gras nachhaltig beeinflusst. Besonders trifft das auf die Huftiere zu. Man unterscheidet dabei Paarhufer (Rinder, Antilopen, Hirsche) und Unpaarhufer (Pferde und Nashörner).
Hornhufe sind das wesentliche Merkmal dieser Tiere. Dabei sind die Zehenspitzen von Hornkapseln umgegeben, die eigentliche Auftrittsfläche ist klein und hart, wodurch sie sich nur wenig abnutzt.
Gräser, Weide- und Raubtiere und Dungkäfer zeigen deutlich verschiedene Nahrungsgewohnheiten. Sie gehören zu Mitgliedern verschiedener Trophieebenen.
Gräser, wie alle anderen grünen Pflanzen auch zählen zu den Produzenten.
Weidetiere sind Konsumenten erster Ordnung oder Primärkonsumenten, sie ernähren sich von den Pflanzen, sind herbivor.
Raubtiere wiederum ernähren sich von den Primärkonsumenten, fressen ausschließlich Fleisch, sind also karnivor und sind Konsumenten zweiter Ordnung oder Sekundärkonsumenten.
Dungkäfer schließlich sind die Zersetzer, deren Nahrungsgrundlage die tote organische Substanz bildet
In einer Nahrungskette sind die einzelnen Trophiestufen miteinanderverbunden, wobei sich der Nachfolger vom Vorgänger ernährt.
Da der Kreislauf durch die Remineralisierung der Destruenten zu den Pflanzen schließt, haben wir wieder den geschlossenen Nahrungskreislauf.
Da die einzelnen Mitglieder der Trophieebenen sich aber nicht ausschließlich von einer Art ernähren, verknüpfen sich Nahrungsketten zu Nahrungsnetzen. Je vielfältiger die Nahrungsgrundlage der einzelnen Arten ist, desto verzweigter sind die Nahrungsnetze. Warzenschweine zum Beispiel sind Allesfresser, sie können tierische wie pflanzlich Nahrung verdauen. Der Schakal ist polyphag, weil er sich sowohl von Aas, Kleintieren als auch von Obst ernähren kann. Der Gepard wird aufgrund dessen, dass er 25 verschiedene Weidetiere jagt, als oligophag bezeichnet Im Gegensatz dazu hat sich das Erdferkel spezialisiert, ausschließlich Ameisen und Termiten werden von ihm gefressen, daher wird es als monophag bezeichnet.
Beschäftigt man sich näher mit dem Nahrungssystem fallen zwei grundlegende Verhältnisse auf:
1. Die Anzahl der Beutetiere ist größer als die Anzahl der Räuber
2. Die Beute wiegt mehr als der Räuber.
Um die Verwertung der Energie bildlich darzustellen, greift man auf die Veranschaulichung in Nahrungspyramiden zurück. Dabei werden die einzelnen Trophiestufen übereinandergestellt, wobei die Basis durch die Produzenten gebildet wird. Die Breite der Stufen wird durch das Gesamtgewicht der Organismen bestimmt. Um einen Einblick in die tatsächlichen quantitativen Verhältnisse zu erhalten, greift man auf die Biomasse der Organismen zurück. Die Biomasse ist das Produkt aus Individuenanzahl und durchschnittliches Gewicht des einzelnen Individuums.
Die Biomasse nimmt dabei mit den steigenden Trophiestufen ab. Eine Faustregel spricht davon, dass die Primärkonsumenten 1/10 der Masse der Produzenten, die Sekundärkonsumenten 1/100 der Primärkonsumenten besitzen. Von 1000kg Gras können sich etwa 100kg Ziegen ernähren, von denen wiederum 10kg Löwen leben. Tatsächlich ist das Verhältnis aber deutlich geringer, da andere limitierenden Faktoren hinzukommen.
Die Anzahl der Individuen und deren Biomasse einer jeden Trophieebene wird durch die zur Verfügung stehende Energie begrenzt.
-
Energiefluss im Ökosystem
Der ökologische Energiefluss hat seinen Ursprung in der Sonne. Er wird für die Lebensgemeinschaft nutzbar, wenn Pflanzen bei der Fotosynthese einen Teil der Lichtenergie einfangen und in chemischen Bindungen festlegen.
Da bei jeder Umformung ein Teil in nicht nutzbare thermische Energie umgewandelt wird - dem System also verloren geht- ist zur Aufrechthaltung der Ordnung des Systems eine permanente Energiezufuhr von außen notwendig. (vgl. 1. und 2. Hauptsatz der Thermodynamik)
Ein anschauliches Bild vom Energiefluss erhält man bei der Betrachtung einer Produktionspyramide. In jeder trophischen Ebene wird ein Teil der Energie eingetragen, der in einer bestimmten Zeiteinheit dazukommt, also die Nettoproduktion pro Zeiteinheit.
Aus einer solchen Pyramide wird die Effizienz des Energieflusses ersichtlich.
Wegen des großen "Energieverlustes" ist die Energieweitergabe als ineffizient zu bezeichnen, die Pyramide wird daher in allen Ökosystemen nach oben hin schmaler.
Interessant ist der Vergleich zwischen Biomasse- und Produktionspyramiden verschiedener Ökosysteme. So ist die Biomasseproduktion der Gräser relativ gering gegenüber der von Gehölzen, die Produktionsmasse fällt jedoch bei ihnen größer aus. Gehölze produzieren nur wenig photosynthetisch aktives Gewebe, der überwiegende Teil hat Stütz- oder Speicherfunktion. Gräser aber bestehen fast ausschließlich aus grünen Organen.
Von allen Lebensräumen des Kontinents hat die Savanne die höchste Sekundärproduktion, ernährt also die meisten Tiere.
In der Savanne leben auf einem ha 150 -200kg Tiere, während in einem ha Wald nur etwa 10kg Tiere leben. Der Grund dafür liegt darin, dass Gräser von den Tieren viel leichter aufgenommen und verwertet werden kann als Holz.
Wegen der hohen Produktivität der Savanne wird diese vom Menschen als Weidegebiet für ihre Herden genutzt und auch häufig Waldgebiete in Grasland (Wiese, Weide oder Getreidefelder) umgebildet. Gefährlich werden diese Handlungen derzeit vor allem dem tropischen Regenwald, der nach und nach gerodet wird, um in kurzzeitig nutzbares Ackerland umgebildet zu werden. Aber auch in der Savanne selbst stehen mittlerweile die Rinderherden mit denen der Wildtiere in starker Konkurrenz. Besonders drastisch fällt dieser Wettbewerb in Afrika aus. Nach regenreichen Perioden und europäischer Unterstützung beim Bau von Brunnen, haben sich die Rinderherden der NomadInnen stark vergrößert, doch jetzt nach jahrzehntelange Dürreperioden hat sich die Situation verschärft und die NomadInnen empfinden die Wildtiere als lästige Konkurrenz, die ihren Herden die Nahrungsgrundlage nimmt. Auch die südamerikanischen Regenwälder müssen dafür sterben, damit sich riesige Rinderherden ausbreiten können, die bei uns bzw. in den USA als Burger landen.
Die Ineffizienz der Energieweitergabe in der Nahrungskette hat auch Folgen für die Ernährungsstrategien der Menschheit. In den reichen Industrieländern des Nordens wird ein großer Teil der pflanzlichen Produktion an die Schlachttiere verfüttert, dabei gehen etwa 90% der gespeicherten Energie verloren. Große Mengen an Kraftfutter werden aus den Entwicklungsländern importiert. Die unter- oder fehlernährte Bevölkerung muss dazu beitragen, uns mit Überschüssen an Butter und Fleisch zu versorgen. Eine überwiegende pflanzliche Ernährung der Menschen würde nicht nur zu einer gesünderen Ernährung führen, sondern auch zu einer geringeren Belastung der Biosphäre und eine bessere Versorgung von mehr Menschen weltweit.
mehr zum Thema Savanne unter:
Etwa 1/3 der Landoberfläche sind Wüsten und ihre Ausbreitung nimmt weiter zu (Deserfikation). In den Subtropen bilden Hitzewüsten einen nahezu geschlossenen Gürtel über die Kontinente der Erde. Die Ausdehnung der Wüste verläuft von vereinzelten kahlen Flecken in der Savanne ausgehend, die sich immer mehr vergrößern und schließlich vereinen. Ursache dafür ist in der Veränderung des Klimas zu sehen, denn seit der letzten Eiszeit verschiebt sich der Wind- und Regengürtel in Richtung Äquator. Seit 3000 trocknen also Seen und Flüsse am Rande der Sahara aus. Hinzu kommt der anthropogene Einfluss. Durch die Überweidung ist der Boden im Umkreis der Brunnen erodiert und die Schaffung immer tieferer Brunnen ließen den Grundwasserspiegel soweit sinken, dass Bäume und Sträucher vertrockneten. (siehe Savanne)
Mittlerweile ist die Ausdehnung soweit vorangeschritten, dass Gebiete zu Wüsten werden, die auch zu Beginn der Eiszeit keine Wüsten gewesen waren. Savannenwälder sterben ab, was durch die Versorgung mit Holz als Brennmaterial der einheimischen Bevölkerung noch beschleunigt wird. Aber während man mittlerweile mehrere Projekte erfolgreich zwischen den Bedürfnissen der EinwohnerInnen und des Waldes vermitteln, schreitet der Raubbau der Industrieländer ungehemmt voran. Rodungsinseln und Transportwege bilden ideale Angriffsflächen, für die Erosion des Boden und damit für die Zerstörung des Regenwaldes.
Das aber hat fatale Folgen für das Klima. Denn die feuchtheißen Luftmassen, die am Äquator aufsteigen kühlen auf dem Weg nach oben ab. Kühlere Luft ist dichter und kann keine Feuchtigkeit bilden. Die Feuchtigkeit kondensiert dabei und fällt als Regen über den Äquatorregionen ab. Ein Teil davon wird durch die Flüssen ins Meer geführt, der Rest verdunstet, bildet kleine Wolken, die sich über benachbarten Gebieten entleeren. Beim Verschwinden des Regenwaldes wird diese Versorgung unterbrochen und die Wüsten breiten sich noch weiter aus.
Die besonderen Merkmale der Wüste in Bezug auf die Lebensbedingungen:
-
Wassermangel, extrem seltene Regenfälle, nur kurze, heftige und unregelmäßige Gewitterschauer
-
große Hitze, die Sonneneinstrahlung wird nicht durch Wolkendecken gemildert, sondern trifft nahezu unvermindert auf der Erdoberfläche auf
-
arides Klima (Verdunstung ist höher als Niederschlagsmenge)
-
große Temperaturunterschiede, die Atmosphäre kann aufgrund der geringen Luftfeuchtigkeit die Wärme nicht speichern, bei Nacht fällt die Temperatur nicht selten um 60°C
-
die trockenen Luft führt zur Verdunstung und dadurch zur Versalzung
-
Nahrungsmangel, wegen der Armut an grünen Pflanzen.
Wasser ist in der Wüster der limitierende Faktor.
Wer in der Wüste überleben will, muss sich an den permanenten Mangel dieses lebensnotwendigen Stoffes gut angepasst haben.
Wasser ist für alle Lebewesen auf verschiedenste Art wichtig. Es dient als
-
Reaktionspartner bei biochemischen Prozessen, wie der Fotosynthese
-
Lösungsmittel bei allen Reaktionen des Stoffwechsels
-
Transportmittel für Ionen und Moleküle (Saftstrom der Bäume, Lymphe und Blut)
-
Kühlmittel, Transpiration ermöglicht Kühlung durch Verdunstung
-
Lebensraum.
Das Gedeihen eines Organismus ist von vielen Umweltfaktoren abhängig. Sein Verschwinden allerdings kann auf das Fehlen eines einzigen Faktors zurückzuführen sein.
Für jedes Lebewesen gibt es hinsichtlich jedes einzelnen Umweltfaktor ein en Toleranzbereich. Innerhalb dessen ist überlebensfähig. Begrenzt wird dieser Toleranzbereich durch ein Minimum und ein Maximum des entsprechenden Faktors. Dort wo das Lebewesen am besten gedeiht liegt sein spezifisches Optimum.
Vorkommen und Häufigkeit einer Art werden durch den Faktor begrenzt, der am weitesten vom Optimum der Art entfernt liegt.
Die Werte sind im einzelnen von Art zu Art verschieden, daher sagt man, dass sich die Arten in ihrer ökologischen Toleranz/ ökologischen Potenz unterscheiden.
Die ökologische Potenz/Toleranz beschreibt die Fähigkeit eines Lebewesens, die gegebenen Umweltbedingungen für sich zu nutzen.
Nach der ökologischen Toleranz unterscheiden sich Xerophyten (Trockenpflanzen), Hyglophyten (Feuchtpflanzen), Hydrophyten (Wasserpflanzen) und Mesophyten (Pflanzen mit keiner ausgeprägten Spezialisierung hinsichtlich des Wasservorkommen).
Worauf es beim Überleben im ariden Klimagebieten wie Wüsten ankommt, ist die Wasserbilanz, also das Verhältnis von aufgenommener und abgegebener Wassermenge. Da in den Dürreperioden ein anhaltendes Wasserdefizit in der Bilanz entsteht, müssen Tiere und Pflanzen und die anderen Organismen sich an solche Trockenzeiten besonders angepasst haben.
Erstaunlich ist die Vielzahl von Anpassungsstrategien, die von den Lebewesen genutzt werden,
Die wichtigsten sind:
-
Widerstand gegen Austrocknung
-
Schutz vor Wasserverlust
-
wasserarme Ausscheidungen (Kot und Urin. )
-
Nahrungs- und Wasserspeicherung
-
Wasserproduktion im Stoffwechsel
Auffällig ist, dass Bewohner der Wüste einen sehr großen Lebensraum beanspruchen, oder anders ausgedrückt, nur wenige Individuen leben hier. Tiere durchstreifen ein sehr großes Territorium, Pflanzen breiten ihre Wurzelsysteme weitflächig aus.
Selbst an den unwirtlichsten Stellen der Wüsten gibt es noch Lebensformen, an die extremen Bedingungen haben sich Flechten angepasst. Die Krustenflechten sind wechselfeucht, d.h. ihr Wasserhaushalt wird durch den Wassergehalt der Umwelt bestimmt. Bei großer Hitze verlieren sie ihr Wasser und ihre Stoffwechselfunktionen werden eingeschränkt bzw. eingestellt, in diesem Zustand überstehen sie auch extreme Hitze. Sie aktivieren ihn wieder, sobald sie durch Nebel oder Tau wieder zu Feuchtigkeit kommen.
Die meisten Pflanzen sterben vor Beginn der Trockenzeit jedoch ab, lediglich ihre Samen bleiben zurück. Diese sind mit harten Schalen ausgestattet, die sie vor der Trockenheit und Hitze schützen, so dass sie Monate oder Jahre zwischen zwei Regenperioden überstehen. Sobald die ersten Regentropfen fallen, können sie binnen weniger Stunden zu keimen beginnen und innerhalb der nächsten 10 - 15 Tage streuen sie ihre Samen aus. Die wohl typischsten Pflanzen der Wüsten und Halbwüsten sind die sogenannten Sukkulenten, die Kakteen. Bei ihnen werden die Anpassungen an den Wassermangel am deutlichsten:
-
flaches weitausbreitendes Wurzelwerk
-
fleischiges, wasserspeicherndes Gewebe
-
dicke, wächserne Cuticula
-
kleine und versenkte Spaltöffnungen, die sich nur nachts öffnen
-
diurnaler Säurerhythmus, CO2 wird nachts aufgenommen und in Form von Apfelsäure gespeichert, die tagsüber wieder abgebaut wird, um CO2 im Calvin-Zyklus (lichtunabhängiger Teil der Fotosynthese) zu verarbeiten
-
Stacheln wehren Tiere ab
-
Stacheln und Haare reflektieren Sonnenlicht und bieten Schutz vor Wasserverdunstung
-
keine Blätter
.Akazien werfen vor der Dürre ihr Blätter ab, während ihre verholzten Teile überdauern können, während bei anderen nur der unterirdische Speicher überlebt.
Dieser hohe Entwicklungsgeschwindigkeit haben sich die Insekten angepasst, denn ihre Eier und Puppen überstehen die Trockenzeit.
Heuschrecken, die verstreut in den Randgebieten der Wüste leben, reagieren auf ausreichende Niederschläge ebenfalls mit sofortiger Eiablage und wachsen in kürzerster Zeit zu riesigen Schwärmen.
Amphibien können diese Zeit durch Ruheperioden überleben. Sie graben sich im Sand ein und verharren dort, wenn sein muss bis zu 10 Monate in tiefer Trockenstarre. Dann nach dem ersten Wolkenbruch kommen sie heraus, paaren sich und legen ihre Eier in Pfützen ab, aus denen dann nach 2-3 Tagen Kaulquappen schlüpfen, die nach ungefähr einem Monat zu Kröten ausgewachsen sind.
Auch andere Tiere ziehen sich unter die Erdoberfläche zurück, die Hornvipern graben sich in den lockeren Sand ein, Wüstenfüchse und Wüstenrennmäuse ziehen sich in ihre Höhlen zurück, in denen ein eigenes Mikroklima mit einer 70% Luftfeuchtigkeit herrscht. Erst wenn bei Nacht die Luft kühler ist beginnen sie ihre Aktivitäten an der Oberfläche.
Bei den Ameisen hingegen werden spezielle Arbeiterinnen mit zuckerhaltigem Saft in Zeiten des Überflusses gefüttert und in Dürrezeiten gemolken.
Eine besondere Form der Anpassung sind bei Wüstenspringmäusen und Kängururatten zu finden. Sie gewinnen die überlebensnotwendige Feuchtigkeit aus ihrer Nahrung, obwohl diese nur aus Samen, also extrem trockener Nahrung besteht. Ihr Trick ist die Nutzung des Oxidationswassers, dass bei der Verwertung der Nahrung entsteht.
(C6H12O6 + 6O2--> 6CO2 + 6 H2O) Mit diesen nur sehr geringen Mengen gehen sie dann äußerst ökonomisch um:
-
ihr Kot ist extrem wasserarm
-
ihr Harn ist hochkonzentriert, so dass bei der Ausscheidung nur wenig Wasser verloren geht
-
Sie haben keine Schweißdrüsen, Abkühlung verschaffen ihnen ihre große Ohren
Eine letzte wichtige Verhaltensweise sei noch erwähnt, nämlich die Wanderung in feuchte Gebiete. Flughühner z.B. legen weite Strecken zurück, um an Wasserstellen zu gelangen, ihr Brustgefieder nimmt zusätzlich noch Wasser auf, mit dem sie ihre Jungen versorgen.
Gebiete der Tundren erstrecken sich nördlich der eurasischen und amerikanischen Waldgrenze und auf einigen antarktischen und arktischen Inseln. Das Klima hier zeichnet sich durch lange, kalte Winter und kurze, kühle Sommer aus. In der langen Polarnacht kommt die Sonne über Monate gar nicht oder nur kaum zum Vorschein, während sie in den Sommermonaten mitunter die ganze Nacht hindurch scheint. Die Vegetationsperiode dauert hier nur etwa 60 Tage.
Weil der Boden auch im Sommer nur etwa 30 -50cm tief auftaut (Permafrost) kann das Wasser nicht versacken, er ist also meist vernässt, weshalb Seen, Sümpfe und Moore das Landschaftsbild prägen.
Bäume sind vorwiegend im südlichen Randgebiet angesiedelt, daran schließt sich die Zone kleiner Sträucher, Gräser an und in ganz nördlichen Regionen herrschen nur noch Flechten, Moose und Algen.
Der limitierende Faktor der Tundra ist eindeutig die fehlende Wärme, die jedes Lebewesen zur Aufrechterhaltung seines Stoffwechsels benötigt. Laut der Reaktionsgeschwindigkeits- Temperatur- Regel erhöht sich die Geschwindigkeit chemischer Reaktionen bei einer Temperaturerhöhung von 10°C um den Faktor 1-3. Die Enzyme der Lebewesen sind auf einen Temperaturbereich von 20-38°C eingerichtet. Nun liegen die Temperaturen der Tundra aber wesentlich unterhalb dieses Optimums, die hier lebenden Organismen müssen sich daher an die extremen Bedingungen angepasst haben.
-
poikilotherme homöotherme Tiere
Größere wechselwarme Tiere sind in der Tundra selten, da der Wärmeverlust durch die Abgabe an die Außenwelt zur Unterkühlung und zum Kältetod führen würde. Es gibt auch fast keine Pflanzenfresser, wie Heuschrecken oder Schnecken, da die Verdauung zu langsam verlaufen würde. Sie sind ausschließlich in den Sommermonaten aktiv und weichen im Winter in wärmere Gebiete aus oder halten Winterruhe bzw. fallen in Kältestarre. Ihre Oberfläche ist meistens dunkel gefärbt, um so mehr Sonnenenergie zu absorbieren.
Zweiflügler - Fliegen und Mücken- sind die häufigsten Insektenarten, in den Sommermonaten wachsen sie zu ungeheueren schwärmen an. Auch sie fallen in Winterruhe, überdauern in der Erde oder als Eier oder Puppen. So überleben sie auch extreme Kälte; ein Laufkäfer aus Alaska übersteht Temperaturen von beinah -90°C.
Eigenwarme Tiere sind von ihrer Umgebungstemperatur nahezu unabhängig, sie halten ihre Körpertemperatur weitgehend konstant. Sie sind auch imstande Pflanzennahrung zu verwerten. Das Aufrechthalten ihrer Körpertemperatur bedarf natürlich immer genügend Energie, denn sobald ihre Körpertemperatur unter 15 -20°C sinkt, tritt der Kältetod ein. Ihre Anpassungsstrategien sehen wie folgt aus:
-
Erhöhen der Wärmeproduktion
-
Tiere der Tundra sind gut isoliert
Bergmannsche Regel: Vögel und Säugetiere kalter Gebiete sind größer als die wärmerer Gegenden, weil ein großes Volumen-Oberflächenverhältnis zu einem geringeren Wärmeverlust führt.
Allensche Regel: Je kühler das Klima, desto kleiner sind die Körperanhänge. Großflächige Körperanhänge (z.B. Ohren, Füße, Schwanz) würden zu großen Wärmeabgaben führen, sind daher eher bei Bewohnern heißer Gebiete anzutreffen.)
Das Fell der hier lebenden Tiere ist sehr dicht, vor allem im Winter, teilweise bedeckt es sogar die Fußsohlen, wie beim Eisfuchs. Darunter sitzt oft eine zentimeterdicke Fettschicht.
-
Verhaltensanpassung wie Abwanderung, Winterschlaf, -ruhe, Aufenthalt im Nest, Einkugeln
Die Lebensgemeinschaft in der Tundra ist einfacher strukturiert als andere und daher für Populationsökologen ein lohnenswertes Objekt. Nur relativ wenige Arten besiedeln das karge Gebiet, insbesondere im Winter, wenn viele Arten gen Süden ziehen. Im Sommer nisten viele sehr viele Vögel hier und es gibt Unmengen von Mücken und Fliegen, aber auch sie sind nur Saisongäste. Die Tundra ist insgesamt also ein sehr inkonstantes Ökosystem.
Je artenärmer eine Gemeinschaft ist und je instabiler die Umwelt, um so größer sind die Schwankungen der Populationsdichte mit der Zeit.
Kurzzeitig können Populationen expotentiell anwachsen.
Lemminge sind allbekannte und liebenswerte Wesen, mit bemerkenswerten Gewohnheiten, sie legen weder Nahrungsvorräte an, noch halten sie Winterschlaf. Sie suchen unter der harten Schneedecke nach Nahrung und vermehren sich auch im Winter. Die Geburtenrate ist sehr hoch, sie werfen bis zu 14 mal in einem Jahr und mit jedem Wurf, werden durchschnittlich 6-8 Lemminge geboren. Die Weibchen werden mitunter schon trächtig, wenn sie selbst noch gesäugt werden. Alle paar Jahre kommt es zu einer regelrechten Massenvermehrung, dann wachsen Lemmingpopulationen expotentiell und alle paar Wochen verdoppelt sich die Zahl der Tiere. Zu Beginn ist noch ausreichend Nahrung und Raum vorhanden ist und die Anzahl der Feinde hält sich in Grenzen. So haben wir also eine hohe Geburtenrate und eine niedrige Sterberate.
Umweltwiderstände begrenzen das Wachstum
Doch meist bricht die expotentiell anwachsende Lemmingpopulation schon innerhalb einer Vermehrungsperiode von einem Jahr zusammen.
Dafür gibt es verschiedene Ursachen:
-
Abwanderung, Lemminge reduzieren ihre Dichte durch Auswanderung. Sobald die Population zu groß wird, ziehen die Tiere in großen Scharen durchs Land. In solchen Jahren findet man sehr häufig Lemmingkadaver außerhalb ihres Verbreitungsgebietes verstreut. Viele ertrinken in Flüssen und Meeresbuchten oder weil sie über Felsenklippen in den Abgrund stürzen. Eine Theorie besagt, dass sie bei ihrer Wanderung immer geradeaus gehen, um nicht wieder zurück ins Ursprungsgebiet zu gelangen. Deshalb folgt eins dem anderen, wenn's sein muss eben auch über Felsenklippen, ins Meer oder in den Fluss.
-
Erhöhung der Sterberate, mit der Zahl der Lemminge nimmt auch die Zahl der Räuber und Parasiten zu, die sich von ihnen ernähren und Krankheiterreger können sich in großen Populationen leichter ausbreiten. Schließlich ist es wohl der Nahrungsmangel durch die Überweidung ihrer Habitate der für die Verringerung der Population sorgt.
-
Verringerung der Geburtenrate, der Dichtestress der Tiere schlägt sich in einem verändertem Hormonhaushalt nieder und dieser bedingt eine Verringerung der Fruchtbarkeit und der Widerstandsfähigkeit gegen Krankheiten.
Jeder Population steht nur ein begrenzter Raum und ein begrenzter Vorrat an Nahrung zur Verfügung, diese und andere limitierenden Faktoren verhindern das Überschreiten der maximalen Bestandgröße. Die Übernutzung eines Lebensraumes lässt ein Population zusammenbrechen. Die Gesamtheit aller Faktoren nennt man Umweltwiderstand.
dichteabhängige / dichteunabhängige Faktoren
Nahrungsmangel, Dichtestress, Feinddruck werden als dichteabhängige Faktoren bezeichnet, da sie auf größere Populationen stärker wirken als auf kleine. Sie können als Regulatoren für die Populationsgröße genannt werden. Ihre Wirkung ist mit der negativen Rückkopplung in Regelkreisen zu vergleichen: je dichter die Population, um so mehr Nahrung wird verbraucht, um so schneller nimmt der Nahrungsvorrat ab und je weniger Nahrung vorhanden ist, um so weniger Lebewesen können sich davon ernähren. Das Prinzip wirkt natürlich in anderer Reihenfolge: je weniger Lebewesen existieren, um so weniger Nahrung wird gebraucht; je mehr übrig bleibt, um so mehr können sich davon ernähren.
Volterraische Regeln
1. Volterrasche Regel: Ernährt sich eine Art von einer anderen, so schwanken die Populationsgrößen von Räuber und Beute periodisch. Die Häufigkeitskurven sind phasenverschoben.
2. Volterraisache Regel: Die Häufigkeit des Räubers und seiner Beute schwanken jeweils um einen Mittelwert.
3.Volterraische Regel: Werden Räuber und Beute gleichermaßen negativ beeinflusst, so nimmt zunächst immer die Zahl der Räuber ab und die der Beute zu.
Diese Regeln stellte der italienische Mathematiker Volterra auf, um die Wechselwirkungen zwischen Beute und Räuber in präziser Form beschreiben zu können. Da sich jedoch ein Räuber nicht ausschließlich von einer Beute ernährt, ist es in der Praxis nicht gewinnbringend mit diesen Regeln zu arbeiten.
Dichteunabhängige Faktoren wirken auf große und kleine Populationsdichten gleichermaßen zu ihnen zählen z.B. die Klimafaktoren.
zyklische Abundanzdynamik (Schwankungen innerhalb der Population)
jährliche Oszillation: innerhalb eines Jahres oder einer Generation regelmäßig stattfindende Veränderung.
mehrjährige Zyklen
Fluktuationen: Häufigkeitsveränderungen zwischen aufeinanderfolgenden Generationen, die meist aus veränderten Umweltbedingungen oder Wanderungserscheinungen resultieren.
Neben unserem Beispiel, den Lemmingen, hat man ebenfalls eine regelmäßige Schwankung der Populationsdichte bei Rot- und Eisfuchs, Bussard, Schneehuhn und Habicht festgestellt. Während bei den Lemmingen die Überweidung ihrer Habitate der ausschlaggebende Grund sein dürfte, warum die Population zusammenbricht, sind Fuchs, Bussard, Habicht und Eulen direkte Folgen der Nahrungsknappheit.
mittlere Dichte einer Population
Die Anpassung an ihre Habitate sorgt dafür, dass die meisten Populationen, bei konstanten Umweltbedingungen, einen relativ konstanten Wert haben.
Bei Wölfe zum Beispiel wird die Bestandsgröße durch ihre Territoriumsgrenzen und die Rudelgröße geregelt. Weil im Rudel sowohl die Geburtenrate als auch die Sterberate dichteabhängig ist, kommt pflanzt sich der Regel nur ein Weibchen fort. Bei zunehmender Größer des Rudels. nimmt die Anzahl der Neugeborenen ab, bei ausbleibender Nahrung kommen gar keine Welpen zur Welt. Die strikte Hierarchie in einem Rudel sorgt dafür, dass die stärksten Tiere stets mit genügend Nahrung versorgt werden, während Außenseiter und Jungtiere leer ausgehen können. Sie müssten das Rudel verlassen , aber dadurch reduzieren sich ihre Chancen noch um ein Vielfaches.
Das Territorium wird gegenüber allen Artgenossen verteidigt, auf einem Gebiet können nie mehr Rudel oder Individuen leben, als das Habitat hergibt, zwischen Fressfeinden oder zwischen Rivalen entstehen Kämpfe um das Revier bzw. das Weibchen, die zum Teil unblutig sein mögen, aber stets einen eindeutigen Ausgang nehmen.
Sobald die optimale Tragfähigkeit des Habitats erreicht ist, hört das Wachstum der Population auf. Die Mortalität ist gleich der Fertalität, die Wachstumsrate also stabil.
Die Wachstumsrate einer Population ist eine genetisch festgelegte Größe, die der Selektion unterliegt. Durch Selektionsvorgänge haben sich im Laufe der Evolution zwei Strategien herausgebildet, auf welche Weise sich eine Population etabliert.
In temporären Habitaten mit stark schwankenden Bedingungen (Wasserpfützen, Überschwemmungsflächen) haben sich Individuen durchgesetzt, die als r-Strategen bezeichnet werden. Unter den kurzzeitig günstigen Bedingungen vermehren sie sich sehr rasch und zeugen sehr viele Nachkommen, die innerhalb kurzer Zeit ihre Entwicklung abgeschlossen haben. Mit dem Verschwinden ihres Lebensraums können sie entweder eine Abwanderung überstehen oder aber sie sind bereits vorher selber wieder zur Fortpflanzung gekommen und hinterlassen nun ihre trockenheitsresistenten Eier.
r- Strategie:
-
große Wachstumsrate
-
kurzlebige Generationen
-
viele Nachkommen
-
schnelle Nutzung der kurzzeitig vorhandenen Ressourcen
-
meist kleine Organismen
-
großes Verbreitungspotential
In stabilen Habitaten mit annährend konstanten Bedingungen, die dauerhaft besiedelbar sind, bringt die ständige Überbelastung der Umwelt keinen Vorteil, die vorhandene Kapazität wird optimal und nicht maximal genutzt. Eine rasche Generationsfolge bringt hier keinen Vorteil, da die knappen Ressourcen noch schneller verbraucht wären. Die Selektion setzt hier bei Genotypen an, deren Träger die Umwelt nicht optimal zu nutzen wissen.
K-Strategie
-
geringe Wachstumsrate
-
langlebige Generationen
-
wenige Nachkommen
-
viele Anpassungen zur effizienten Nutzung der Ressourcen
-
meist große Organismen
-
geringes Verbreitungspotenzial
-
elterliche Fürsorge (bei Tieren)
![]()